
LE MUTAZIONI DEL DIAMANTE MANDARINO
(Taeniopygia guttata castanotis)
(Scheda riassuntiva ed informativa sulla specie presente in Allevamento Nevada; testi e fotografie potrebbero avere fonti varie e molteplici, di libera consultazione)
Il Diamante mandarino, originario dell’Australia, è sicuramente l’uccello esotico più allevato e conosciuto nel mondo.
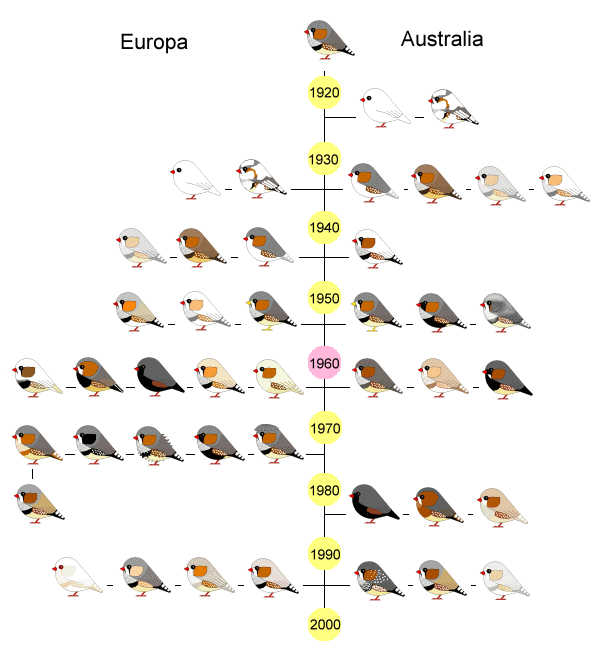
In cattività questo piccolo uccello è ormai allevato e selezionato con innumerevoli “forme” e “colori”; le prime differenze che si possono notare guardando fotografie su libri o in internet sono infatti la differenza della taglia e l’innumerevole varietà di mutazioni che lo riguardano.
In tanti anni di selezione la taglia di un Diamante Mandarino di forma ancestrale (selvatica) risulta nettamente inferiore rispetto al Diamante Mandarino allevato a scopi espositivi secondo un preciso standard di razza.
Lo standard, infatti, prevede certi canoni a cui avvicinarsi (riporteremo a fine discussione alcuni standard in vigore); la grandezza (quindi la taglia) è uno di questi.
Normalmente i Diamanti Mandarino selezionati vengono chiamati Maggiorati (per ovvi motivi) o Olandesi. È infatti da attribuire agli allevatori dell’Olanda il merito del lavoro selettivo più grosso e quindi la scelta dei canoni da seguire per lo standard, attualmente in vigore anche in Italia.
In Inghilterra (per fare un esempio) avremo invece dei Diamanti Mandarino “inglesi”; secondo il loro standard la grandezza della testa deve essere decisamente maggiore. Per questo motivo ci ritroveremo davanti sempre lo stesso animale ma con un “capoccione” di notevoli dimensioni.
(stupenda immagine illustrativa di http://www.everyoneweb.com/worldwidezeb/ )

Queste grandi differenze spesso traggono in inganno i neofiti che si ritrovano realmente davanti animali molti diversi tra loro; difficile credere che appartengono sempre alla stessa specie.
Discorso a parte merita il “Diamante Mandarino di Timor” (Taeniopygia guttata guttata), una sottospecie del Taeniopygia guttata da non confondere assolutamente per un Diamante Mandarino non selezionato o di taglia piccola.
Tornando al nostro caro Diamante Mandarino (Taeniopygia guttata castanotis) è bene sottolineare che la forma ancestrale continua ad essere sempre presente presso qualsiasi negozio ed allevamento, a differenza del “Diamante Mandarino Selezionato / Maggiorato / Olandese”, ovviamente più ricercato.
Prendendo spunto da alcune foto del caro amico Stefano Giannetti, vi proponiamo le due “forme” a confronto; nella prima foto si può notare un ancestrale Grigio (piccolo) ed un Grigio Dorso Chiaro. Nella seconda foto due mutati Bruni (quello di destra è anche portatore di Petto Arancio). Oltre alla struttura e alla taglia si può notare anche come la selezione si sia spinta sulla colorazione di base.


La forma selvatica ed ancestrale è quindi quella piccola di color GRIGIO e spesso si presenta con l’iride dell’occhio rosso (carattere tramandato dai soggetti selvatici).
Molti soggetti (anche di varie mutazioni) che provengono da ceppi NON realmente selezionati (quindi dalla classica forma “piccola”) possono presentare anch’essi l’iride di colore rosso, sempre derivato dalle caratteristiche genetiche della forma selvatica. Il Diamante Mandarino selvatico a livello oculare presenta pochissima melanina (da qui deriva il colore rosso dell’occhio) e per questo motivo possiamo ritrovare questa particolare caratteristica (che ben differisce dal concetto di mutazione) tramandata anche a molti nostri “amici di casa”.
In casi eccezionali è possibile osservare soggetti in natura di color Bruno. Infatti la mutazione Bruno è stata osservata in soggetti selvatici catturati nel 1927 in Australia per poi esser fissata in cattività.
Per questo motivo il GRIGIO ed il BRUNO sono mutazioni che definiamo MUTAZIONI BASE. Tutte le altre mutazioni si sovrappongo a queste due BASI creando quindi tutte le varietà di colore che ad oggi possiamo notare.
Prima di vedere le mutazioni più comini procediamo ad un breve glossario di genetica (utile per capir la trasmissione delle mutazioni stesse):
FENOTIPO = è ciò che appare all’esterno; l’aspetto… ciò che vedete ad occhio nudo.
GENOTIPO = è la costituzione genetica, l’insieme di geni localizzati nei cromosomi.
Gene DOMINANTE = è un gene che negli accoppiamenti di prima generazione predomina sul gene alternativo (recessivo), apparendo sempre nel fenotipo. Basta quindi un solo gene mutato per trasmettere interamente la mutazione.
Gene RECESSIVO = è un gene che tende a mantenersi latente (nascosto) negli individui eterozigoti. Pertanto nel fenotipo compare solo il gene dominante, a meno che il gene recessivo non sia presente in doppia dose (omozigosi).
AUTOSOMICO = aggettivo che si riferisce agli autosomi, cioè a cromosomi che non partecipano alla determinazione del sesso. È l’opposto di “sessolegato”, quindi indica un carattere non legato al sesso dell’individuo.
SESSOLEGATO = significa che un dato carattere è legato al sesso dell’individuo. È l’opposto di “autosomico” e indica che il gene in questione è situato su uno dei cromosomi sessuali. **
** Negli uccelli il maschio possiede una coppia uguale di cromosomi sessuali ZZ , mentre la femmina ZW. Usiamo la lettera Z al posto della lettera X e la lettera W al posto della lettera Y perché i cromosomi sessuali XY vengono utilizzati per l’essere umano che, al contrario rispetto agli uccelli, l’uomo ha come cromosomi sessuali la coppia XY e la donna la coppia uguale XX
Semplificando: Nella prole, il maschio trasmette quindi sempre un cromosoma sessuale Z; se la femmina a sua volta trasmette il cromosoma Z nasceranno maschi (ZZ), se la femmina trasmette il cromosoma W nasceranno femmine (ZW).
Tutte le mutazioni sessolegate conosciute ad oggisi collocano sul cromosoma Z. Per questo motivo i maschi possono essere interamente MUTATI se presentano quella mutazione in doppia dose (Z “mutata” + Z “mutata”) oppure portatori di tale mutazioni se presentano un solo cromosoma mutato (Z “mutata” + Z).
Nel Fenotipo un portatore di mutazione sessolegata (o recessiva) è quasi uguale ad un esemplare non mutato (salvo alcune piccole e quasi impercettibili caratteristiche).
Una femmina, invece, avendo un solo cromosoma sessuale Z può essere Mutata direttamente (Z “mutata” + W) oppure NON mutata (Z + W). Non esistono (per ovvi motivi) femmine portatrici di mutazioni sessolegate.
Eumelanine: melanine corrispondenti al colore nero/bruno.
Feomelanine: melanine di colore rosso/bruno.
Useremo alcune stupende foto di Stefano Giannetti fatte in occasione della famosa mostra Zebra’s per il “Zebrafinken Club Italia” e “sfrutteremo” spunti presi da alcuni siti a tema di grandissimo interesse (come Mr Mandarino, e-finch, ecc…).
In qualità di socio del club stesso sicuramente non faremo torti a nessuno utilizzando questo materiale in forma prettamente divulgativa!

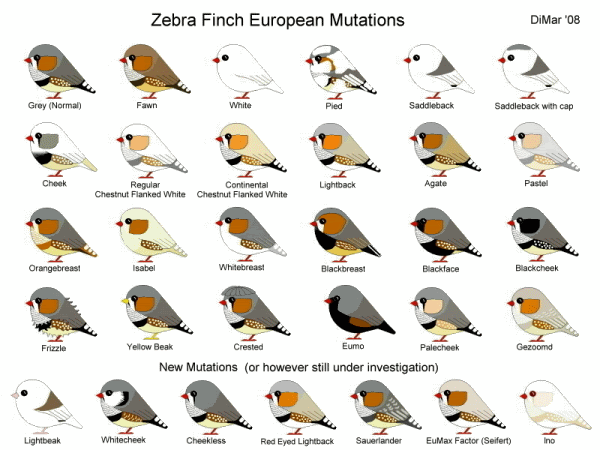
LE BASI
GRIGIO
(mutazione Base, è la variante in cattività della forma selvatica)
Il Grigio è la prima mutazione BASE; presenta la massima ossidazione della eumelanina e la minima quantità di feomelanina. I segni distintivi (lacrima, coda, barra pettorale) sono neri, mentre il ventre risulta color crema quasi bianco. Il dorso è privo di sfumature brune.


BRUNO
(mutazione Sessolegata)
La mutazione Bruno è da considerarsi la seconda mutazione BASE. È apparsa in soggetti selvatici catturati nel 1927 in Australia per poi esser fissata in cattività. Le eumelanine non sono completamente ossidate e la feomelanina rimane invariata. Il color grigio diventa quindi bruno ed il nero non ha la possibilità di manifestarsi interamente.


LE MUTAZIONI PIU’ COMUNI
Ogni mutazione si presenta in maniera FENOTIPICA diversa su base GRIGIA e su base BRUNA; quasi tutte le mutazioni possono anche “sommarsi” tra loro ottenendo quindi soggetti plurimutati (fatta eccezione per le mutazioni sessolegate).
DORSO CHIARO (DC)
(mutazione Sessolegata; Allelica Dominante rispetto al MON e al MOR)
Parziale riduzione di entrambe le melanine (sia eumelanine che feomelanine). Siccome nella testa e nella nuca si trovano molte eumelanine e sul dorso si presenta prettamente la feomelanina, si nota molto lo stacco di colore fra la testa ed il dorso stesso. Il ventre risulta bianco candido. La mutazione è apparsa in Germania nei primi anni 50; inizialmente confusa con la mutazione pastello venne scartata e successivamente ripresa (e fissata) in Svizzera.



MASCHERATO OCCHI NERI (MON)
(mutazione Sessolegata; Allelica Recessiva al Dorso Chiaro, Allelica Dominante al MOR e al Lutino)
Questa mutazione, apparsa in Australia nel 1937, riduce in maniera molto forte l’eumelanina e la feomelanina. Molto somigliante alla MOR non viene presentata in nessuna esposizione proprio perché un soggetto MON è uguale (ma molto più “slavato”) ad un soggetto MOR. I Fianchi e la Guancia (nei maschi) e la coda (anche nelle femmine) risulta di un tenue color sabbia. Il resto del corpo color bianco/crema. I Soggetti ancestrali (non di selezione) presentano spesso la testa (soprattutto negli esemplari femmina) più scura e punteggiata di grigio.
N.B. il colore degli occhi non è vincolante e non “c’entra nulla”! È “solo” il nome della mutazione! Soggetti adulti e selezionati presentano quindi l’iride dello stesso colore delle altre mutazioni.


MASCHERATO OCCHI ROSSI (MOR)
(mutazione Sessolegata; Allelica Recessiva al Dorso Chiaro)
Questa mutazione è apparsa in Europa nei primi anni ‘70, la sua origine però è dubbia perché spesso confusa con la mutazione Mascherato Occhi Neri. Nella testa, il dorso, le ali ed il ventre, notiamo una forte riduzione di eumelanina e di feomelanina. Le melanine dei segni distintivi però si riducono solo parzialmente. Guance color arancio, fianchi marroni e soprattutto coda Molto Scura (segno di maggior distinzione tra MON e MOR) sono le caratteristiche di questa mutazione.
N.B. Come per il MON, il colore degli occhi non è vincolante e non “c’entra nulla”! È “solo” il nome della mutazione! I MOR da pulli nascono con l’occhio effettivamente di colore rosso, crescendo perdono questa particolarità ed i soggetti adulti e selezionati presentano quindi l’iride dello stesso colore delle altre mutazioni.



FACCIA NERA (FN)
(mutazione Dominante, non è possibile quindi avere soggetti portatori)
Mutazione apparsa nel 1959 in Germania. L’eumelanina è più marcata; l’area tra la striscia del becco e quella dell’occhio si fondono formando un unico disegno triangolare; la barra pettorale si espande sul ventre scurendolo, quindi “trasformandolo” in nero.


GUANCIA NERA (GN)
(mutazione Autosomica – Recessiva)
Questa mutazione, scoperta in Germania nel 1977, colpisce le eumelanine; la guancia risulta di colore nero (è presente sia nei maschi che nelle femmine) ed il fianco del maschio da marrone diventa nero. Spesso si possono notare soggetti di poca selezione che presentano il difetto tipico nei fianchi: guancia color nero e fianco marrone o nero sporco di marrone.


GUANCIA
(mutazione Dominante, non è possibile quindi avere soggetti portatori, in doppia dose è letale)
Apparsa in Europa nei primi anni ‘70 (in Belgio), spesso viene confusa con la mutazione Mascherano combinata alla mutazione Guancia Nera. La forte diminuzione delle melanine sul mantello e nei segni distintivi ne modifica quindi il disegno, ottenendo così un soggetto interamente bianco/crema, senza disegno nella coda e con guancia e fianchi color bruno scuro. Anche le femmine presentano la guancia. La mutazione è ancora considerata “particolare” perché spesso può generare problemi visivi nei soggetti. Questi problemi variano dalla comparsa di bolle sugli occhi fino alla totale assenza del bulbo oculare nei nascituri.


FEOMELANICO (FEO) (ex ISABELLA)
(mutazione Autosomica – Recessiva)
Questa mutazione, apparsa in Olanda nei primi anni ’60, causa una riduzione totale delle eumelanine; il piumaggio è costituito quindi da sole feomelanine. Da qui appunto il nome FEOMELANICO. Il corpo risulta interamente color crema nei soggetti a base Bruna, grigio chiaro nei soggetti a base Grigia. L’intensità dei disegni varia lasciando visibili ma scoloriti i caratteristici quadri della coda.


PETTO NERO (PN)
(mutazione Autosomica – Recessiva)
Apparsa in Germania nella prima metà degli anni ’60, questa mutazione è il risultato dell’aumento generale della feomelanina. Nel maschio la guancia si allunga verso la nuca ed il collo; scompare interamente la lacrima. Gli scacchi della coda lasciano il posto a piume a base bianca, i pallini nel fianco del maschio si ovalizzano, scompare la zebratura del petto che si fonde con la barra pettorale, le remiganti e le copritrici alari diventano orlate di bianco ed arancio. Nella femmina si può notare l’assenza della lacrima ed il particolare della coda (come nel maschio).





PETTO ARANCIO (PA)
(mutazione Autosomica – Recessiva)
La mutazione PETTO ARANCIO agisce sull’eumelanina, che viene sostituita interamente dalla feomelanina. Tutti i caratteri neri diventano quindi marroni. La lacrima ed i mustacchi spariscono (sia nei maschi che nelle femmine) e la coda diventa quindi a scacchi bianchi ed arancioni/marroni.



PETTO BIANCO (PB) (ex PINGUINO)
(mutazione Autosomica – Recessiva)
Apparsa nei primi anni ‘30 in Australia, la mutazione PETTO BIANCO si identifica dalla diminuzione della eumelanina e della feomelanina. I disegni distintivi, sia nel maschio che della femmina (mustacchi, lacrima, barra pettorale, zebratura) spariscono del tutto, ad eccezione dei quadri alternati della coda. Nella femmina la guancia risulta bianca, nel maschio guancia e fianchi rimangono invariati dalla forma base.



BIANCO
(mutazione Autosomica – Recessiva)
La mutazione BIANCO è la prima mutazione del Diamante Mandarino comparsa (Danimarca 1927). L’intero corpo diventa bianco candido perché tutte le melanine non si manifestano. Siccome tutti i caratteristici segni distintivi spariscono, anche differenziare un maschio da una femmina risulta molto problematico. A livello fenotipico l’unico modo per distinguere quindi i sessi è la colorazione del becco; nel maschio il rosso risulta più acceso. I Portatori di Bianco possono essere confusi con Pezzati o con portatori di Pezzato perché i soggetti si presenteranno Grigi o Bruni (a seconda della base) con piume e penne bianche (molto pezzati, poco pezzati o con poche zone bianche; una semplice remigante o il sottogola). Tecnicamente a determinare il fenotipo bianco sono responsabili due geni, indicati con i nomi “pezzato dominante” e “pezzato recessivo“.
Un bianco, geneticamente parlando, è obbligatoriamente omozigote per il gene pezzato recessivo, mentre il gene pezzato dominante può essere presente sia in omozigosi che in eterozigosi.
Per ottenere un soggetto bianco quindi i genitori dovranno essere necessariamente entrambi almeno portatori del gene pezzato recessivo e almeno uno dei due deve presentare il gene pezzato dominante quanto meno in singola dose.


PEZZATO
(mutazione Autosomica – Recessiva)
Strettamente legata alla mutazione BIANCO (Danimarca 1927), le melanine si comportano nello stesso modo ma solo in alcune parti del piumaggio. Le zone interamente bianche si alternano a quelle colorate. Nella riproduzione dei Bianchi non è raro ottenere soggetti pezzati perché identificati come portatori di tal mutazione. “Ovviamente” un soggetto pezzato portatore di Bianco avrà nel suo genotipo il gene del Bianco e potrà generare figli interamente candidi; un “semplice” pezzato invece difficilmente genererà soggetti tutti Bianchi (pezzato “tutto” pezzato).


SCUDATO
(mutazione Autosomica – Recessiva)
Selezionata a partire dal 1974/1975 in Olanda, la mutazione SCUDATO è in realtà una particolare selezione della mutazione PEZZATO. Tutto il corpo si presenta bianco tranne il dorso ed il codione che mantengono il colore di base del soggetto. Data la difficoltà nella sua selezione si può presentare anche la “variante” con calotta; la nuca rimane quindi dello stesso colore dello scudo.


* NOTA BENE: Anche se l’articolo è molto riassuntivo vogliamo ricordare che: “Il fattore per il bianco e quello per le pezzature sono due fattori che agiscono sugli stessi geni ma hanno due meccanismi completamente diversi…”
PASTELLO
(mutazione suddivisa in mutazione “PASTELLO DOMINANTE” e mutazione “PASTELLO RECESSIVA”)
La mutazione PASTELLO viene fissata in Australia nei primi anni ’30. La forte riduzione delle melanine genera soggetti con tutto il piumaggio generale molto chiaro, color argenteo. La guancia nei maschi, quindi, risulta beige. Ci sono due forme di pastello; una geneticamente Dominante ed una geneticamente Recessiva.




Altre mutazioni presenti nel Diamante Mandarino sono: CIUFFATO, BECCO GIALLO, LUTINO, EUMO, ecc… ecc…

SCHEDE TECNICHE
Riportiamo a seguire le Schede Tecniche Ufficiali 2008 / 2009 / 2010 / 2011
(Redatte dalla Commissione Tecnica Nazionale I.E.I. – Ficeti, Caimi, Piccinini)
1) Grigio, Bruno, Dorso Chiaro, Mascherato, Bruno Mascherato, Bruno Dorso Chiaro
2) Petto Nero: Grigio, Bruno, Dorso Chiaro, Mascherato, Bruno Mascherato, Bruno Dorso Chiaro
3) Faccia Nera: Grigio, Bruno, dorso Chiaro, Mascherato
4) Guancia Nera: Grigio, Bruno, Dorso Chiaro, Mascherato
5) Petto Arancio: Grigio, Bruno, Dorso Chiaro, Mascherato, Bruno Mascherato, Bruno Dorso Chiaro
6) Diluito: Grigio, Bruno, Dorso Chiaro, Bruno Dorso Chiaro
7) Petto Bianco: Grigio, Bruno
10) Topazio: Grigio, bruno, Dorso Chiaro, Bruno Dorso Chiaro
12) Bianco/Scudato: Grigio, Bruno